第七章 怎样识别杏仁
如果你是一个徒步旅行者,吃腻了农场上种植的瓜果蔬菜,那么试一试吃一点野生食物,就是一件有趣的事。你知道,有些野生植物,如野草莓和乌饭树的蓝色浆果,不但味道好,而且吃起来安全。它们的样子同我们所熟悉的农家品种相当类似,所以我们能够很容易地把这些野生浆果认出来,虽然它们比我们种植的那些品种要小得多。爱冒险的徒步旅行者在吃蘑菇时小心翼翼,因为他们知道蘑菇中有许多品种吃了会致人死命。但即使是最爱吃干果的人也不会去吃野生的杏仁,因为其中有几十种含有足以致人死命的氰化物(即纳粹毒气室中使用的那种毒物)。森林中到处都有其他许多被认为不能吃的植物。
然而,所有的作物都来自野生植物。某些野生植物是怎样变成作物的呢?这是一个特别令人困惑的问题,因为许多作物(如巴旦杏)的祖先不是吃了叫人送命就是味道糟得难以入口,而其他一些作物(如玉米)在外观上同它们的野生祖先又差别太大。住在岩洞里的那些男女是些什么样的人,竟会想出“驯化”植物这个主意来?而这又是如何做到的呢?
植物驯化可以定义为:栽种某一植物并由此有意或无意地使其发生不同于其野生祖先的、更有利于人类消费的遗传变化。对作物的培育在今天是一种由专业科学家去做的、自觉的、高度专业化的工作。他们已经了解现存的数以百计的作物,并着手开发新的品种。为了实现这一目标,他们种下了许多种子或根,选出最好的后代,再种下它们的种子,把遗传学知识用来培育能够繁殖纯种的优良品种,也许甚至会利用最新的遗传工程技术来转移某些有用的基因。加利福尼亚大学戴维斯分校有一个系(果树栽培学系),全系专门研究苹果,还有一个系(葡萄栽培与葡萄酒工艺学系),专门研究葡萄和葡萄酒。
但是,植物驯化的历史可以追溯到1万多年前。最早的农民当然不会利用分子遗传技术来得到这种结果。这些最早的农民甚至没有任何现存的作物可以作为样品来启发他们去培育新的作物品种。因此,他们不可能知道,不管他们做什么,他们最终都能一饱口福。
那么,最初的农民是怎样在不知不觉中驯化了植物的呢?比方说,他们是怎样在不知道自己在做什么的情况下把有毒的杏仁变成了无毒的杏仁的呢?除了使某些野生植物变得更大或毒性更少外,他们实际上还使它们产生了哪些变化呢?即使是一些重要的作物,驯化的时间也大不相同:例如,豌豆驯化的时间不迟于公元前8000年,橄榄在公元前4000年左右,草莓要等到中世纪,美洲山核桃则更迟至1846年。许多能出产食物的重要的野生植物为千百万人所珍视,例如在世界上的许多地方,人们为了得到可吃的橡实而寻找橡树,但橡树甚至在今天也仍然没有驯化。是什么使得某些植物比其他植物更容易驯化,或更吸引人去对其驯化?为什么橄榄树向石器时代的农民屈服了,而橡树则仍然使我们最聪明的农艺师一筹莫展呢?
让我们首先从植物的观点来看一看驯化的问题。就植物来说,我们不过是无意间“驯化”了植物的成千上万种的动物之一。
和其他所有动物(包括人)一样,植物必须将其子代向它们能够茁壮生长的地区扩散,并传递其亲代的基因。小动物的扩散靠行走或飞翔,但植物没有这样的选择自由,因此它们必须以某种方式沿途搭便车旅行。有几种植物的种子生得便于被风吹走或在水上漂流,而其他许多植物则把种子包在好吃的果子里并用颜色和气味来宣告果子的成熟,从而哄骗动物把种子带走。饥饿的动物把这果子扯下来吃掉,或者走开,或者飞走,然后在远离亲代树的某个地方把种子吐出或随粪便排出。种子可以用这种办法被带到几千英里之外。
植物的种子连你的肠胃都无法消化,而且仍然能够从你的排泄物中发出芽来,了解到这一点,你也许会感到惊奇。但任何爱冒险的而又不太容易呕吐的读者不妨做一做这方面的试验,亲自来验证一下。许多野生植物的种子实际上必须通过动物的肠胃,然后才能发芽。例如,非洲有一种瓜很容易被一种形似鬣狗的叫做土狼的动物吃掉,结果这种瓜大多数都生长在土狼排泄的地方。
关于想要沿途搭便车旅行的植物是怎样吸引动物的这个问题,可以考虑一下野草莓的例子。当草莓的种子还没有成熟、不能马上播种时,周围的果实是又青又酸又硬。当种子最后成熟时,果实就变得又红又甜又嫩。果实颜色的变化成了一种信号,把鸫一类的鸟儿吸引来啄食果实然后飞走,最后吐出或随粪便排泄出种子。
当然,草莓并不是有意要在种子准备好撒播的时候去招引鸟儿。鸫也不是有意要去驯化草莓。相反,草莓是通过自然选择来演化的。未成熟草莓的颜色越青和味道越酸,在种子成熟前来吃草莓从而使种子遭到破坏的鸟儿就越少;成熟草莓的味道越甜和颜色越红,来撒播成熟种子的鸟儿就越多。
其他无数的植物都有适合于让某些种类的动物吃并撒播的果实。正如草莓适合于鸟儿一样,橡实适合于松鼠,芒果适合于蝙蝠,某些莎草适合于蚂蚁。这符合我们关于植物驯化的部分定义,因为祖代植物在遗传方面的变化使它更有利于人类消费。但没有人会一本正经地把这种演化过程称为驯化,因为鸟儿、蝙蝠和其他动物不符合那个定义的另一部分:它们不是有意去栽种植物的。同样,作物从野生植物开始演化的早期无意识阶段包括这样的一些演化方式,即植物吸引人类去采食并撒播它们的果实,但还不是有意去栽种它们。人类的排泄处所和土狼的一样,也许就是最早的无意识的作物培育者的一个试验场。
我们在许多地方偶然地播下了我们所吃的植物的种子,我们的排泄处仅仅是其中之一。当我们采集到可吃的野生植物并把它们带回家的时候,有些在路上或家里散落了。有些水果在种子还仍然十分完好的时候就腐烂了,于是就被扔进了垃圾堆,不再吃它。草莓的种子是果实的一部分,实际上也被我们送入口中,但由于种子太小,最后还是被吞了下去,并随粪便排了出来。但还有些水果的种子相当大,就被吐了出来。因此,我们的茅坑加上我们的痰盂和垃圾堆一起构成了最好的农业研究实验室。
不管这些种子最后到了哪个“实验室”,它们都是来自某种可吃的植物——即我们出于某种原因喜欢吃的植物。从你采摘浆果的那些日子起,你就知道挑选某些浆果或浆果灌木。最后,当最初的农民开始有意识地去播种时,他们播下的必然是他们存心采集的那些植物的种子,虽然他们还不懂大浆果的种子有望长成能够结出更多大浆果的灌木这一遗传原则。
因此,当你在一个炎热、潮湿的日子里艰难地走进到处都是蚊子的多刺的灌木丛中时,你并非只是为了任何一丛草莓才这样去做的。即使是无意识的,你还是决定了哪一丛草莓看上去最有希望,以及它是否值得你来这一趟。你的潜意识中的判断标准是什么?
一个判断标准当然就是大小。你喜欢大的浆果,因为你不值得为几颗难看的小浆果去被太阳晒烤和蚊子叮咬。这就是许多作物的果实比它们野生祖先的果实大得多的部分原因。超市里的草莓和乌饭树的蓝色浆果同野生的品种相比显得硕大肥壮,这情形是我们特别熟悉的;这方面的差异也只是最近几百年才出现的。
在其他植物方面的这种大小差异,可以追溯到农业出现的初期,那时人工栽培的豌豆经过筛选和演化,其重量超过野生豌豆的10倍。狩猎采集族群在几千年里采集的就是这种小小的野生豌豆,就像我们今天采集乌饭树的蓝色浆果一样。然后,他们才有选择地去收获和种植那些最吸引人的最大的野生豌豆——这就是我们所说的农业——它开始自动地促使豌豆的平均大小每一代都有增加。同样,超市里的苹果的直径一般在3英寸左右,而野生苹果的直径只有1英寸。最早的玉米棒子的长度几乎不超过半英寸,但公元1500年墨西哥印第安农民已经培育出长达6英寸的玉米棒子,而现代的玉米棒子则可长达一英尺半。
我们所栽种的植物的种子和它们的许多野生祖先的种子的另一个明显的差异是在味苦方面。许多野生植物的种子为防动物把它们吃掉,在演化过程中味道变苦而难以入口,或竟然有毒。因此,自然选择对种子和果实所起的作用正好相反。果实好吃的植物让动物来撒播它们的种子,但果实里的种子必须很难吃。否则,动物就会把种子嚼碎,种子也就发不出芽来了。
关于苦味的种子及其在驯化过程中的变化,杏仁提供了一个引人注目的例子。大多数野巴旦杏的种子都含有一种叫做苦杏仁苷的其苦无比的化学物质,这种物质(前面已提到过)在分解时产生了有毒的氰化物。如果有人竟会蠢到不顾野生杏仁苦味的警告而去吃它,那他就会因此而丧命。既然无意识驯化的第一阶段是采集可以吃的种子,那么对野生杏仁的驯化又是怎样达到这第一阶段的呢?
对此的解释是:偶然有几棵巴旦杏树在一个基因上产生了突变,使它们不能合成苦味的苦杏仁苷。这些树在荒野里灭绝了,没有留下任何后代,因为鸟儿发现了它们的种子,把种子吃个精光。但最初的农民的孩子们由于好奇或饥饿,把他们周围的野生植物每一种都弄一点来尝尝,最后竟品尝到并发现了这些没有苦味的巴旦杏树。(同样,如果今天的欧洲农民偶然发现了几棵橡实甜而不苦的橡树,他们仍然会感到庆幸。)这些不苦的巴旦杏的种子就是古代农民可能会去种的种子,开头是无心地任其生长在垃圾堆上,后来则是有意地种在自己的果园里了。
不迟于公元前8000年的野生杏仁,已在发掘出来的希腊考古遗址中出现。到公元前3000年,在地中海以东的土地上已有野生杏仁在驯化。埃及国王图坦卡蒙[1]于公元前1325年左右去世后,放在他的著名陵墓里供他死后享用的食品中就有杏仁。在其他许多为人们所熟悉的作物中有利马豆、西瓜、马铃薯、茄子和卷心菜。这些作物的野生祖先,有的味苦,有的有毒,其中偶然也有几个味道甘美的品种,但那必定是从古代旅行者的排泄处长出来的。
如果说大小和味道是狩猎采集族群选择野生植物的最明显的标准,那么其他标准则包括果实肉多或无子、种子含油多以及纤维长。野南瓜种子外面的果肉很少,或根本没有果肉,但早期的农民喜欢选择果肉比种子多得多的南瓜。很久以前,人们在栽培香蕉时就选择了全是肉而没有种子的品种,从而启发了现代农业科学家去培育无籽柑橘、无籽葡萄和无籽西瓜。水果无籽是一个很好的例子,说明人类的选择是怎样把野生水果原来的演化作用完全颠倒了过来,因为原来的演化作用实际上只是撒播种子的一种手段。
在古代,有许多植物都是由于含油的果实和种子而同样得到选择的。在地中海地区最早得到驯化的果树有油橄榄,栽培的时间大约在公元前4000年以后,栽培的目的就是为了得到它的油。人工栽培的油橄榄比野生的油橄榄不但果实大,而且含油率也高。古代农民为了得到含油的种子而选择了芝麻、芥菜、罂粟和亚麻,而现代的植物学家为了同样的目的而选择了向日葵、红花和棉花。
为了油而培育棉花,这还是近代的事。在这之前,当然是为了纺织用的纤维而选择棉花。棉花的纤维(或称棉绒)是棉花种子上的茸毛,美洲和旧大陆的早期农民为了得到长的棉绒独立地选择了不同品种的棉花。另外还有两种作为古代纺织品原料而栽种的植物是亚麻和大麻,它们的纤维来自茎,因此对它们的选择标准是又长又直的茎。虽然我们把大多数作物看作是为吃而种植的,但亚麻也是我们最早的作物之一(驯化时间不迟于公元前7000年)。它是亚麻布的原料,它一直是欧洲主要的纺织原料,直到工业革命后为棉花和合成纤维所代替。
迄今为止,我们对野生植物演化为作物所描述的种种变化,与早期农民实际上可能已注意到的一些特征直接有关——如果实的大小、苦味、多肉、含油和纤维的长度。通过收获这些具有特别可取的品质的野生植物,古代人在无意识中传播了这些植物,使它们走上了驯化之路。
然而,除此以外,至少还有其他4种主要变化未能引起采摘浆果的人去作出引人注目的选择。在这些情况下,如果采摘浆果的人的确引起了什么变化,那或是由于其他植物因某些不明原因始终无法得到而收获了可以得到的植物,或是由于改变了对植物起作用的选择条件。
第一个这样的改变影响了种子传播的野生结构。许多植物都有其传播种子的专门机制(从而使人类无法对它们进行有效的采集)。只有由突变产生的缺乏这种结构的种子才会被人收获而成为作物的祖先。
一个明显的例子就是豌豆。豌豆的种子(我们吃的豌豆)封闭在豆荚里。野豌豆要发芽生长,就必须破荚而出。为了做到这一点,豌豆就演化出一种基因,能使豆荚突然破裂,把豌豆弹射到地上。偶然产生突变的豌豆的豆荚不会爆裂。在野外,这种产生突变的豌豆一直到死都是包裹在亲株上的豆荚里面,而只有这种能爆裂的豆荚才把它们的基因传递给后代。但与此相反的是,人类能够收获到的唯一豆荚可能就是留在植株上的那些不爆裂的豆荚。因此,一旦人类开始把野豌豆带回家去吃,立刻就产生了对这种单基因突变的选择。对同样不爆裂突变进行选择的例子还有兵豆、亚麻和罂粟。
野生的小麦和大麦不是封闭在能够爆裂的荚子里,而是长在麦秆的顶端,麦秆能自动脱落,使种子落到能够发芽生长的地面上。一种单基因突变使麦秆不会脱落。在野生状态下,这种突变对植物来说可能是毁灭性的,因为种子不能落地,就无法发芽生根。但这些产生突变的种子省力地留在秆子上可能是在等待人类来收获它们并把它们带回家。当人类接着种下了这些收获来的产生突变的种子后,农民又一次可以从这些种子的后代中得到所有产生突变的种子,收获它们,播种它们,而后代中未产生突变的那些种子就落到地上,而无法得到了。这样,农民就180度地改变了自然选择的方向:原来成功的基因突然变得具有毁灭性了,而毁灭性的突变却变得成功了。1万多年前,这种对不脱落的小麦和大麦麦秆的无意识的选择,显然是人类对植物的第一个重大的“改良”。这个变化标志着新月沃地农业的开始。
第二个改变甚至是古代旅行者更难以觉察的。对于在气候变化无常的地区生长的一年生植物来说,如果所有的种子都迅速地同时发芽,那可能是毁灭性的。如果发生了这种情况,那么只要一次干旱或霜冻就可能把幼苗全都杀死,连传种接代的种子都没有了。因此,许多一年生植物演化的结果是通过发芽抑制剂来减少损失,使种子在开始阶段休眠,然后在几年里分批发芽。这样,即使大多数幼苗由于一次恶劣的天气而死光,剩下的一些种子还会在以后发芽。
野生植物赖以减少损失的一种普遍的适应性变化,是把它们的种子裹在一层厚厚的皮壳或保护层中。产生这种适应性变化的这许多植物包括小麦、大麦、亚麻或向日葵。虽然这些后发芽的种子仍然有机会在野外发芽,但请考虑一下随着农业的发展而必然发生的情况。早期的农民可能在反复试验中发现,他们可以通过松土、浇水然后播种的办法来获得更高的产量。如果是这样,种子就会立即发芽并长成植物,它们的种子又可以收获下来在来年种下。但许多野生植物的种子不会立即发芽,因此种下去也不会有任何收成。
野生植物中偶然产生突变的个体没有厚厚的种皮,也没有其他的发芽抑制剂。所有这些突变体迅速发芽,最后产生了突变的种子。早期的农民可能没有注意到这种差异,他们只知道去注意和有选择地收获大的浆果。但播种—生长—收获—播种这种循环会立即无意识地选择了那些突变体。和种子传播方面的变化一样,发芽抑制方面的这些变化是小麦、大麦、豌豆以及其他许多作物的共同特点,而这是它们的野生祖先所没有的。
早期农民觉察不到的另一个重要变化与植物的繁殖直接有关。植物培育的一个普遍问题是:偶然产生突变的植物个体比正常的个体更有益于人类(例如,由于种子较大和苦味较少)。如果这些可取的突变体接下去同正常的植物进行品种间的杂交,这种突变就会立刻被淡化或完全失去。那么,对早期农民来说,在什么情况下这种突变会继续保留下来?
对自我繁殖的植物来说,突变体会自动地保留下来。对无性繁殖(用亲代植物的块茎或根来繁殖)的植物或能够自花受精的雌雄同株的植物来说,情况也是如此。但大多数野生植物都不是用这种方法来繁殖的。它们或者是雌雄同株,但不能自花受精,必须同其他雌雄同株的个体进行品种间的杂交(我的雄蕊使你的雌蕊受精,你的雄蕊使我的雌蕊受精),或者像所有正常的哺乳动物一样,是雌雄异体。前一种植物叫做自交不亲和雌雄同株,后一种植物叫做雌雄异株。这两种情况对古代农民来说都不是好消息,因为他们可能因此而糊里糊涂地失去所有对他们有利的突变体。
解决办法涉及另一种难以觉察的变化。许多植物的突变影响到其自身的繁殖系统。有些产生突变的个体甚至不需要授粉就可以结出果实来,其结果就使我们有了无籽的香蕉、葡萄、柑橘和菠萝。有些产生突变的雌雄同株植物失去了它们的自交不亲和性,而变得能够自花受精——许多果树,如李、桃、苹果、杏和樱桃就是这一过程的例证。有些产生突变的葡萄通常都是雌雄异体,但也会变成自花受精的雌雄同株。古代的农民虽然还不懂得植物繁殖生物学,但仍然用所有这些办法最后得到了一些有用的作物,这些作物能够繁殖纯种,因而值得去重新种植,而不是那些本来被看好的突变体,它们的后代则因为毫无价值而湮没无闻。
因此,农民就是从一些特有的植物中进行选择,他们选择所根据的标准,不但有大小和味道这些看得见的品质,而且还有诸如种子传播机制、发芽抑制和繁殖生物学这些看不见的特点。结果,不同的植物由于十分不同的或甚至相反的特点而得到选择。有些植物(如向日葵)由于大得多的种子而得到选择,而另一些植物(如香蕉)则由于种子小或没有种子而得到选择。选择生菜则取其茂盛的叶子而舍其种子或果实;选择小麦和向日葵则取其种子而舍其叶;选择南瓜则取其果实而舍其叶。特别有意思的是,由于不同的目的,对于一种野生植物可以有不同的选择,从而产生了外观十分不同的作物。甜菜在巴比伦时代即已有种植,不过那时种植甜菜是为了它们的叶(如叫做牛皮菜的现代甜菜品种),后来再种植则是为了它们可吃的根,最后(在18世纪)则是为了它们所含的糖份(糖用甜菜)。最早的卷心菜可能原本是为了它们的含油种子而被种植的,后来经过甚至更大的分化,对它们也就有了不同的选择:有的选择了叶(现代的卷心菜和羽衣甘蓝),有的选择了茎(撇蓝),有的选择了芽(抱子甘蓝),有的选择了花芽(花椰菜和花茎甘蓝)。
迄今为止,我们已经讨论了野生植物由于农民有意无意的选择而变成了作物的问题。就是说,农民开始时选择了某些植物个体的种子,带回来种在自家的园子里,然后每年都挑选一些后代的种子,再在下一年种在园子里。但这种变化的很大一部分也由于植物的自我选择而受到了影响。达尔文所说的“自然选择”指的是一个物种的某些个体在自然条件下比同一物种中与之竞争的个体可以生存得更好与(或)繁殖得更成功。实际上,是差别生存状况和繁殖状况的自然过程作出了这种选择。如果条件改变了,不同种类的个体有可能生存或繁殖得更好,从而“被自然所选择”,其结果就是这个种群经历了演化改变。一个典型的例子是英国飞蛾工业黑化现象的发展:在19世纪,随着环境变得更脏,颜色深的飞蛾比颜色浅的飞蛾更为普通,因为栖息在深暗肮脏的树上的深色飞蛾,相形之下比浅色飞蛾更有可能逃脱捕食者的注意。
与工业革命改变了飞蛾的环境几乎一样,农业耕作也改变了植物的环境。经过松土、施肥、浇水和除草的园地所提供的生长环境,完全不同于山坡上干燥、未施肥的园地。植物在驯化中所产生的许多变化都来自此类条件的变化,因此也就有了那些处于有利条件的植物品种的变化。例如,如果农民在园子里播下的种子密密麻麻,那么在种子之间就会产生剧烈的竞争。大的种子可以利用良好的条件迅速生长,此时它们所处的地位就比小的种子有利,因为小的种子先前是长在干燥、未施肥的山坡上的,而那里种子比较稀少,竞争也不那么激剧,所以只有生长在那样的地方才对它们有利。植物本身之间这种持续的竞争,对于获得较大的种子和野生植物变成古代作物期间逐步形成的其他许多变化,起到了重要的促进作用。
就植物的驯化来说,有些植物在很久以前就驯化了,有些直到中世纪才驯化,而还有一些野生植物竟然不受我们所有活动的影响。这方面的巨大差异的原因是什么?我们可以考察一下不同作物在西南亚新月沃地发展的固定顺序来推断出许多答案。
事情原来是这样的:新月沃地最早的作物,如小麦、大麦和豌豆,大约是在1万年前驯化的,它们都起源于呈现许多优势的野生祖先。它们已经可以食用,而且在野生状态下产量很高。它们很容易生长,只要播种或栽植就行了。它们生长迅速,播种后不消几个月就可收获了。对于仍然处于流浪的猎人和定居的村民之间的早期农民来说,这是一个巨大的优点。它们很容易贮藏,这和后来的许多作物如草莓和生菜不同。它们大都是自花传粉:就是说,各种作物都是自己给自己传授花粉,把它们自己的合意的基因毫无改变地传递下去,而不必同其他的对人类不大有用的品种进行杂交。最后,为了转化为作物,它们的野生祖先很少需要在基因方面产生什么变化——例如,就小麦来说,只要产生使麦粒不脱落和迅速而均匀的发芽的突变就行了。
作物培育的下一阶段包括在公元前4000年左右驯化的最早的果树和坚果树,其中有橄榄树、无花果树、枣树、石榴树和葡萄藤。同谷物和豆科植物相比,它们的缺点是种植后至少要3年才开始结实,而达到盛产期则要等到10年之后。因此,只有那些已完全过上了定居的乡村生活的人,才有可能种植这些作物。然而,这些早期的果树和坚果树仍然是最容易栽种的作物。和后来驯化的树木不同,它们可以用插枝甚或播种的办法来直接栽种。插枝还有一个好处:一旦古代农民发现或培育了一棵多产的果树,他们可以确信,这棵树的所有后代可以长得和它一模一样。
第三阶段涉及一些栽培起来难得多的果树,包括苹果、梨、李和樱桃。这些树不能靠插枝来种植。用种子来种也是白费力气,因为即使是其中优秀品种的后代也十分易变,所结的果实也多半没有价值。这些树要靠困难的嫁接技术来种植,而这个技术在中国农业开始后很久才发展起来。即使你懂得嫁接的原理,嫁接也仍然是一项困难的工作。不仅如此,这原理本身也只有通过有意识的实验才能发现。发明嫁接这种事情,大概不再是什么一个流浪者在某个排泄处方便,后来在返回时惊喜地发现那里竟因此而长出了结有甘甜水果的树来。
许多在晚近阶段发展起来的果树提出了另外一个问题,因为它们的野生祖先完全不是自花传粉。它们必须由属于同一品种但产生遗传变异的另一植物进行异花传粉。因此,早期的农民要么必须去找到不需要异花传粉的果树,要么必须有意识地去种植不同的遗传品种或同一果园中附近的雄性和雌性个体。所有这些问题把苹果、梨、李和樱桃的驯化时间一直推迟到古典时期前后。然而,大约与此同时,还有一批在晚近出现的驯化植物却得来全不费工夫,它们本来都是长在有意栽培的作物地里的杂草之类的野生植物。原本是杂草的作物包括黑麦、燕麦、芜菁、萝卜、甜菜、韭葱和生菜。
虽然我刚才详细介绍的一系列作物适用于新月沃地,但部分类似的一系列作物也出现在世界其他地方。尤其是,新月沃地的小麦和大麦是被称之为谷物(属禾本科)的那类作物的代表,而新月沃地的豌豆和兵豆则是豆类(属豆科,包括大豆)的代表。谷类作物的优点是生长快,碳水化合物含量高,每公顷耕地可产1吨食物。因此,今天的谷物占人类消耗的全部卡路里的半数以上,并包括现代世界上12种主要作物中的5种(小麦、玉米、稻米、大麦和高粱)。许多谷类作物蛋白质含量低,但这一缺陷可以由豆类来弥补,因为豆类的蛋白质通常达25%(大豆为38%)。因此,谷物和豆类一起为均衡饮食提供了许多必不可少的成分。
正如表7.1扼要说明的那样,当地谷物和豆类组合的驯化,标志着许多地区粮食生产的开始。最为人熟知的例子,是新月沃地的小麦和大麦与豌豆和兵豆的组合,中美洲的玉米与几种豆类的组合,以及中国的稻米和小米与大豆和其他豆类的组合。不大为人所知的是非洲高粱、非洲稻米和珍珠稗与豇豆和野豆的组合,以及安第斯山脉的非谷类的昆诺阿藜与几种豆类的组合。
表7.1同时表明,为获得纤维在新月沃地曾对亚麻进行过早期驯化,在其他地方也有类似情况。大麻、4种棉花、丝兰和龙舌兰在不同时期为中国、中美洲、印度、埃塞俄比亚、非洲撒哈拉沙漠以南地区和南美洲提供了制绳和织布的纤维,在这些地区的几个地方,还用驯化动物的绒毛作为补充。在早期粮食生产的这些中心中,只有美国东部和新几内亚仍然没有纤维作物。
同这些类似之处相比,全世界的粮食生产体系中也存在着某些重大的差异。其中的一个差异是:在世界上的许多地方,农业开始涉及种子撒播和农田单作以及最后用牲畜犁地等问题。就是说,用手把种子一把把撒下去,从而使整块田因而只种一种作物。一旦牛、马和其他大型哺乳动物得到驯化,它们就被套上了犁,于是农田就由畜力来耕作。然而,在新大陆还没有驯化过任何可以套上犁头的动物。相反,在那里耕地始终是用手持的棍棒或锄头,而种子也是用手一颗颗种下去,而不是满把的撒播。因此,新大陆的大部分园地都是许多作物混种在一起,而不是单作。
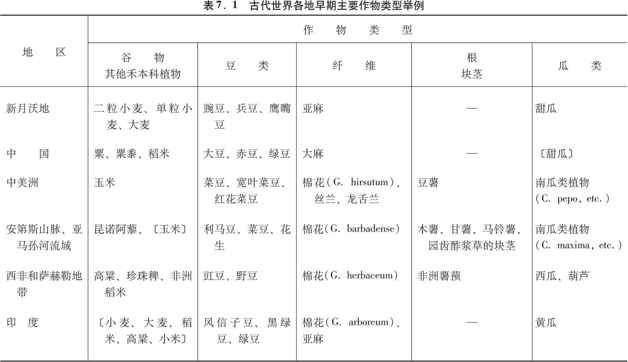
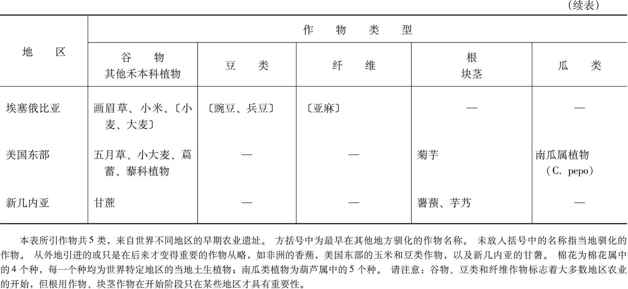
农业体系中的另一个重大差异涉及卡路里和碳水化合物的主要来源问题。我们已经看到,在许多地区,这方面的主要来源是谷物。不过,在另一些地区,谷物的这一任务被根和块茎接管了或分担了,虽然根和块茎在古代的新月沃地和中国是无关紧要的。在赤道南美洲主食是木薯和甘薯,在安第斯山脉是马铃薯和圆齿酢浆草的块茎,在非洲是非洲薯蓣,在东南亚和新几内亚是印度洋-太平洋地区的薯蓣和芋艿。树生作物主要的有香蕉和面包果,它们也是东南亚和新几内亚的富含碳水化合物的主食。
因此,到了罗马时代,今天的几乎所有作物都已在世界上的某个地方得到驯化。正如我们还将在家畜方面看到的那样(第九章),古代的狩猎采集族群非常熟悉当地的野生植物,而古代的农民显然也已发现并驯化了几乎所有值得驯化的动物。当然,中古的僧侣确已开始栽培草莓和树莓,而现代的培育植物的人仍在改良古代的作物,并已增加了一些新的次要作物,主要是一些浆果(如乌饭树的蓝色浆果、越橘和猕猴桃)和坚果(澳洲坚果、美洲山核桃和腰果)。但和古代的一些主食如小麦、玉米和稻米相比,这些新添的现代作物始终只具有不太大的重要性。
不过,在我们所列举的关于驯化成功的例子中仍然缺乏许多野生植物。尽管它们具有食用价值,但我们却不曾成功地驯化它们。在我们驯化失败的这些例子中,引人注目的是橡树。橡实不但是欧洲农民在荒年作物歉收时的应急食物,而且也是加利福尼亚和美国东部的印第安人的主食。橡实具有营养价值,含有丰富的淀粉和油。和许多在其他方面可以食用的野生食物一样,大多数橡实含有味苦的丹宁酸,但爱吃橡实的人学会了用处理杏仁和其他野生植物中味苦的化学物质的同样办法来处理丹宁酸:或者用研磨和过滤来去掉丹宁酸,或者只从丹宁酸含量低的偶然产生突变的橡树上收获橡实。
为什么我们不能驯化像橡实这样宝贵的粮食来源呢?为什么我们花了那么长的时间去驯化草莓和树莓?对那些植物的驯化,即使是掌握了像嫁接这样困难的技术的古代农民也会束手无策,这又是怎么一回事呢?
事情原来是这样的:橡树有3个不利因素。首先,它们生长缓慢,可能使大多数农民失去耐心。小麦种下去不消几个月就可得到收成;杏仁种下去3、4年后就可长成能够结果实的树;但种下一颗橡实可能在10年或更长的时间里不会有什么收益。其次,橡树所结的坚果无论是大小还是味道都适于松鼠,而我们都见到过松鼠埋藏、挖掘和吃橡实的情景。如果偶尔有一颗橡实松鼠忘记把它挖出,那么这颗橡实就可长出橡树来。有数以10亿计的松鼠,每一只松鼠每一年把数以百计的橡实传播到几乎任何一个适于橡树生长的地方。这样,我们人类就不可能为我们所需要的橡实去选择橡树。橡树生长缓慢和松鼠行动迅速这些问题大概也说明了为什么山毛榉和山核桃树同样未能驯化的原因,虽然欧洲人和美洲土著分别对这两种树种大量地加以利用以获得它们的坚果。
最后,杏仁和橡实的最重要差异也许是:杏仁的苦味由单一的优势基因所控制,而橡实的苦味似乎由许多基因所控制。如果古代农民栽种了偶然产生不苦的突变的杏仁或橡实,那么根据遗传规律,如果是巴旦杏树,结果,长成的树上的杏仁有一半可能也是不苦的,而如果是橡树,则几乎所有橡实可能仍然是苦的。仅仅这一点就足以使任何想要种橡实的农民的热情荡然无存,尽管他们已经赶走了松鼠并且保持耐心。
至于草莓和树莓,我们在与鸫和其他喜吃浆果的鸟儿的竞争中遇到了同样的困难。是的,罗马人的确在他们的园子里照料过野草莓。但是,由于千百万只欧洲鸫把野草莓的种子排泄在每一个可能的地方(包括罗马人的园子里),所以草莓始终是鸫想要吃的小浆果,而不是人想要吃的大浆果。由于近来保护网和温室的发展,我们才终于能够把鸫打败,并根据我们自己的标准来重新设计草莓和树莓。
因此,我们已经看到,超市上的大草莓和野生的小草莓之间的差异只是一个例子,用来说明把人工培育的植物与其野生祖先区别开来的许多特征。这些差异首先来自野生植物本身之间的自然变异。有些变异,如浆果的大小和坚果的苦味方面的变异,可能很快就被古代的农民注意到了。其他变异,如种子传播机制或种子休眠方面的变异,在现代植物学兴起之前,可能并未被人类认出来。但是,不管古代旅行者对可食用的野生植物的选择是否依赖于自觉的或不自觉的选择标准,由此而产生的野生植物向作物的演化起先总是一种无意识的过程。这是我们对野生植物个体进行选择的必然结果,是园子里各植物个体之间竞争的结果,而这种竞争所偏爱的个体和在野外得天独厚的个体是不同的。
这就是为什么达尔文在他的伟大著作《物种起源》中并不是一开始就解释自然选择问题的原因。他的第一章反而详细说明了我们的驯化动植物是如何通过人类的人为选择而出现的。达尔文不是讨论我们通常认为和他联系在一起的加拉帕戈斯群岛[2]上的鸟类,而是一上来就讨论——农民是怎样培育出不同品种的醋栗的!他写道,“我已经看到园艺学著作中对园丁们在用这样差的材料取得这样了不起的成果方面的令人叹为观止的技术所表现出来的巨大的惊奇;但这种技术是简单的,就其最后结果来说,对这一技术的采用也几乎是无意识的。它在于总是去培育最出名的品种,播下它的种子,然后当碰巧出现了一个稍好一点的品种时,再去选择它,就这样地进行下去。”通过人为选择来培育作物的这些原则仍然可以成为我们的关于物种起源通过自然选择的最可理解的模式。
注 释: